The monofunctional mitochondrial and peroxisomalenzymes are found in the mitochondria and peroxisomes of eukaryotes, respectively. The multifunctional enzymes are found in bacteria and in the peroxisomes of some eukaryotes, but they serve two functions: the N-terminal domain works the same as the other classes of enoyl-CoA isomerases and the C-terminal domain works as a dehydrogenase, specifically, to 3-hydroxyactyl-CoA.[4] There are two divisions among the mitochondrial enoyl Co-A isomerase: short-chain and long-chain [4].[6] In an immunoblot, antibodies were run against all enoyl CoA isomerase. However, two of these isomerases had antibody attachment: the short chain isomerase and the peroxisomal multifunctional enzyme.[6] There was one enzyme which did not have binding specificity to this antibody: mitochondrial long-chain isomerase. Long-chain isomerase was found when it eluted at a lower potassium phosphate concentration in the gradient.[6][7] Thus, the discovery of three sub-classes of enoyl CoA isomerase was made.
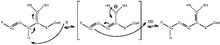
If the enzyme is unclear, react it with an enoyl CoA Derivative. If the enzyme gives off more than one product, it is a multifunctional enzyme. If it gives off one product, it is solely enoyl Co-Al isomerase.[10]
Mitochondria (both short- and long-chain) of rat liver contain more than one enoyl Co-A isomerase.[10] To further support the idea that short- and long-chain isomerases elute at different concentration of potassium phosphate concentration, they do not share similar primary polypeptide structure, hence they must not be evolutionarily related.[6][11]Peroxisomes of plants and of rat liver are very different in the way they operate. Despite their primary structure similarities, there are differences among the different specimen. To begin with, the peroxisomes of rat liver are a multifunctional enzyme including enoyl-CoA isomerase, enoyl-CoA hydratase, and L-(−)-3-hydroxyacyl-CoA dehydrogenase.[12] Three different enzymes reside on this entity (multifunctional protein) allowing this enzyme to perform isomerization, hydration, and dehydration.[13][14] Isomerase activity on the multifunctional enzyme occurs at the amino-terminal catalytic half of the protein along with the hydratase activity.[15] The dehydrogenase activity of enoyl-CoA occurs in the carboxyl-terminal.[15] Upon further investigation of the CoA binding site on the amino-terminal half of the multifunctional protein, the CoA substrate is not transferred through the aqueous phase from the isomerization phase to the site of hydration or does not have a bulk phase.[11][16] This removes the need for a substrate transferring enzyme.[17] On the other hand, the cotyledons convert long-chain 3-trans-enoyl-CoA, long-chain 3-cis-enoyl-CoA, and short-chain 3-cis-enoyl-CoA species into their 2- trans-enoyl-CoA respective forms.[13] As previously mentioned, plant enoyl-CoA isomerase exclusively forms the 2-trans isomer as product. It does not act on 4-cis-enoyl-CoA species or 2-trans- 4-trans-dienoyl-CoA species.[13] In comparing the products of the plant peroxisome and the multifunctional enzyme of rat liver, the plant has no hydratase activity.[13] The Plant form did not form a 2-cis-isomer (from enoyl-CoA hydratase) or D- or L- 3 hydroxy derivative (L-(−)-3-hydroxyacyl-CoA dehydrogenase): products of multifunctional enzyme of rat liver.[13] The turnover rates of these the two sub divisions of peroxisomes are very different. The Kcat/Km ratio in cotyledons is 10^6 M-1s-1 which outperforms the ratio .07 * 10^6 M-1s-1.[13] Due to a high turnover rate, the plant peroxisomes contain a lesser amount of enoyl-CoA isomerase than their counterparts in the rat liver.[13]
In rat liver, mitochondrial enoyl CoA isomerase and peroxisomal enoyl CoA isomerase embedded in the multifunctional enzyme have similarities in the primary structure sequence.[15] When comparing the amino-terminal half of E. coli against the amino-terminal half of rat liver, there were primary and secondary structure similarities towards the middle of the amino-terminal end.[15] This conserved region must be important for structure and function of this specific enzyme since showing equally in both E. coli and rat liver.[15][18]
More recent studies link hepatitis C virus (HCV) infection to defects in fatty acid degradation, specifically, to that in enoyl-CoA isomerase.[23]HCV is the leading cause of chronic hepatitis, cirrhosis, and liver cancer, and more than 180 million people are affected globally.[24] Due to the prolonged latency of the virus and no existing cures to rid the virus specifically,[25]HCV is a serious problem that is causing more deaths than HIV/AIDS in the United States,[26] but its threat still do not receive adequate attention. The need for a HCV-specific treatment is essential, and according to John Ward, the director of the CDC Hepatitis Division, it can save up to 120,000 lives.[26]
^ abPartanen ST, Novikov DK, Popov AN, Mursula AM, Hiltunen JK, Wierenga RK (September 2004). "The 1.3 A crystal structure of human mitochondrial Delta3-Delta2-enoyl-CoA isomerase shows a novel mode of binding for the fatty acyl group". J. Mol. Biol. 342 (4): 1197–208. doi:10.1016/j.jmb.2004.07.039. PMID15351645.
^Patricia C. Babbitt; George L. Kenyon (1992). "Ancestry of the 4-Chlorobenzoate Dehalogenase: Analysis of Amino Acid Sequence Identities among Families of Acyl: Adenyl Ligases, Enoyl-CoA Hydratases/Isomerases, and Acyl-CoA Thioesterases". Biochemistry. 31 (24): 5594–5604. doi:10.1021/bi00139a024. PMID1351742.
^Anu M. Mursula; Daan M. F. van Aalten; J. Kalervo Hiltunen; Rik K. Wierenga (2001). "The Crystal Structure of delta3-delta2-Enoyl-CoA Isomerase". Molecular Biology. 309 (4): 845–853. doi:10.1006/jmbi.2001.4671. PMID11399063. S2CID69172923.
^Stoffel W, Grol M (December 1978). "Purification and properties of 3-cis-2-trans-enoyl-CoA isomerase (dodecenoyl-CoA delta-isomerase) from rat liver mitochondria". Hoppe-Seyler's Z. Physiol. Chem. 359 (12): 1777–82. doi:10.1515/bchm2.1978.359.2.1777. PMID738702.