Nicotinamide adenine dinucleotide phosphate, abbreviated NADP+ or, in older notation, TPN (triphosphopyridine nucleotide), is a cofactor used in anabolic reactions, such as the Calvin cycle and lipid and nucleic acid syntheses, which require NADPH as a reducing agent ('hydrogen source'). NADPH is the reduced form, whereas NADP+ is the oxidized form. NADP+ is used by all forms of cellular life.[1]
NADP+ differs from NAD+ by the presence of an additional phosphate group on the 2' position of the ribose ring that carries the adeninemoiety. This extra phosphate is added by NAD+ kinase and removed by NADP+ phosphatase.[2]
Biosynthesis
NADP+
In general, NADP+ is synthesized before NADPH is. Such a reaction usually starts with NAD+ from either the de-novo or the salvage pathway, with NAD+ kinase adding the extra phosphate group. ADP-ribosyl cyclase allows for synthesis from nicotinamide in the salvage pathway, and NADP+ phosphatase can convert NADPH back to NADH to maintain a balance.[1] Some forms of the NAD+ kinase, notably the one in mitochondria, can also accept NADH to turn it directly into NADPH.[3][4] The prokaryotic pathway is less well understood, but with all the similar proteins the process should work in a similar way.[1]
NADPH
NADPH is produced from NADP+. The major source of NADPH in animals and other non-photosynthetic organisms is the pentose phosphate pathway, by glucose-6-phosphate dehydrogenase (G6PDH) in the first step. The pentose phosphate pathway also produces pentose, another important part of NAD(P)H, from glucose. Some bacteria also use G6PDH for the Entner–Doudoroff pathway, but NADPH production remains the same.[1]
Ferredoxin–NADP+ reductase, present in all domains of life, is a major source of NADPH in photosynthetic organisms including plants and cyanobacteria. It appears in the last step of the electron chain of the light reactions of photosynthesis. It is used as reducing power for the biosynthetic reactions in the Calvin cycle to assimilate carbon dioxide and help turn the carbon dioxide into glucose. It has functions in accepting electrons in other non-photosynthetic pathways as well: it is needed in the reduction of nitrate into ammonia for plant assimilation in nitrogen cycle and in the production of oils.[1]
There are several other lesser-known mechanisms of generating NADPH, all of which depend on the presence of mitochondria in eukaryotes. The key enzymes in these carbon-metabolism-related processes are NADP-linked isoforms of malic enzyme, isocitrate dehydrogenase (IDH), and glutamate dehydrogenase. In these reactions, NADP+ acts like NAD+ in other enzymes as an oxidizing agent.[5] The isocitrate dehydrogenase mechanism appears to be the major source of NADPH in fat and possibly also liver cells.[6] These processes are also found in bacteria. Bacteria can also use a NADP-dependent glyceraldehyde 3-phosphate dehydrogenase for the same purpose. Like the pentose phosphate pathway, these pathways are related to parts of glycolysis.[1] Another carbon metabolism-related pathway involved in the generation of NADPH is the mitochondrial folate cycle, which uses principally serine as a source of one-carbon units to sustain nucleotide synthesis and redox homeostasis in mitochondria. Mitochondrial folate cycle has been recently suggested as the principal contributor to NADPH generation in mitochondria of cancer cells.[7]
NADPH can also be generated through pathways unrelated to carbon metabolism. The ferredoxin reductase is such an example. Nicotinamide nucleotide transhydrogenase transfers the hydrogen between NAD(P)H and NAD(P)+, and is found in eukaryotic mitochondria and many bacteria. There are versions that depend on a proton gradient to work and ones that do not. Some anaerobic organisms use NADP+-linked hydrogenase, ripping a hydride from hydrogen gas to produce a proton and NADPH.[1]
Like NADH, NADPH is fluorescent. NADPH in aqueous solution excited at the nicotinamide absorbance of ~335 nm (near UV) has a fluorescence emission which peaks at 445-460 nm (violet to blue). NADP+ has no appreciable fluorescence.[8]
NADH and NADPH are very stable in basic solutions, but NAD+ and NADP+ are degraded in basic solutions into a fluorescent product that can be used conveniently for quantitation. Conversely, NADPH and NADH are degraded by acidic solutions while NAD+/NADP+ are fairly stable to acid.[12]
Enzymes that use NADP(H) as a coenzyme
Adrenodoxin reductase: This enzyme is present ubiquitously in most organisms.[13] It transfers two electrons from NADPH to FAD. In vertebrates, it serves as the first enzyme in the chain of mitochondrial P450 systems that synthesize steroid hormones.[14]
Enzymes that use NADP(H) as a substrate
In 2018 and 2019, the first two reports of enzymes that catalyze the removal of the 2' phosphate of NADP(H) in eukaryotes emerged. First the cytoplasmic protein MESH1 (Q8N4P3),[15] then the mitochondrial protein nocturnin[16][17] were reported. Of note, the structures and NADPH binding of MESH1 (5VXA) and nocturnin (6NF0) are not related.
^Iwahashi Y, Hitoshio A, Tajima N, Nakamura T (April 1989). "Characterization of NADH kinase from Saccharomyces cerevisiae". Journal of Biochemistry. 105 (4): 588–93. doi:10.1093/oxfordjournals.jbchem.a122709. PMID2547755.
^Iwahashi Y, Nakamura T (June 1989). "Localization of the NADH kinase in the inner membrane of yeast mitochondria". Journal of Biochemistry. 105 (6): 916–21. doi:10.1093/oxfordjournals.jbchem.a122779. PMID2549021.
^Hanukoglu I, Rapoport R (Feb–May 1995). "Routes and regulation of NADPH production in steroidogenic mitochondria". Endocrine Research. 21 (1–2): 231–41. doi:10.3109/07435809509030439. PMID7588385.
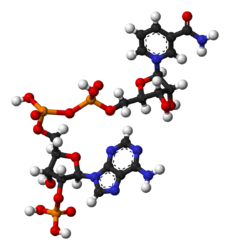 NADP+
NADP+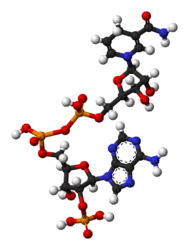 NADPH
NADPH

