-
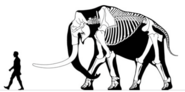 Mammuthus meridionalis bull, around 4 metres (13 ft) tall
Mammuthus meridionalis bull, around 4 metres (13 ft) tall -
 Steppe mammoth (M. trogontherii) around 3.9 metres (13 ft) tall in front-on (without head) side-on and top-down views
Steppe mammoth (M. trogontherii) around 3.9 metres (13 ft) tall in front-on (without head) side-on and top-down views -
 Columbian mammoth (M. columbi) bull around 3.7 metres (12 ft) tall
Columbian mammoth (M. columbi) bull around 3.7 metres (12 ft) tall -
 Woolly mammoths (M. primigenius), including one of the largest, the Siegsdorf mammoth (left, around 3.5 metres (11 ft) tall), and a mature Siberian bull (around 2.7 metres (8.9 ft) metres tall)
Woolly mammoths (M. primigenius), including one of the largest, the Siegsdorf mammoth (left, around 3.5 metres (11 ft) tall), and a mature Siberian bull (around 2.7 metres (8.9 ft) metres tall)
| Mammoth | |
|---|---|

| |
| Columbian mammoth in the Page Museum in Los Angeles. | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Proboscidea |
| Family: | Elephantidae |
| Subfamily: | Elephantinae |
| Tribe: | Elephantini |
| Genus: | †Mammuthus Brookes, 1828 |
| Type species | |
| †Elephas primigenius[1][2] Blumenbach, 1799
| |
| Species | |
| Synonyms | |
| |
A mammoth is any species of the extinct elephantid genus Mammuthus. They lived from the late Miocene epoch (from around 6.2 million years ago) into the Holocene about 4,000 years ago, and various species existed in Africa, Europe, Asia, and North America. Mammoths are distinguished from living elephants by their (typically large) spirally twisted tusks and in at least some later species, the development of numerous adaptions to living in cold environments, including a thick layer of fur.
Mammoths and Asian elephants are more closely related to each other than either of them are to African elephants. The oldest mammoth representative, Mammuthus subplanifrons, appeared around 6 million years ago during the late Miocene in what is now southern and Eastern Africa.[3] Later in the Pliocene, by about three million years ago, mammoths dispersed into Eurasia, eventually covering most of Eurasia before migrating into North America around 1.5–1.3 million years ago, becoming ancestral to the Columbian mammoth (M. columbi). The woolly mammoth (M. primigenius) evolved about 700–400,000 years ago in Siberia, with some surviving on Russia's Wrangel Island in the Arctic Ocean until as recently as 4,000 years ago, still extant during the existence of the earliest civilisations in ancient Egypt and Mesopotamia.
Etymology and early observations
According to The American Heritage Dictionary, the word "mammoth" likely originates from *mān-oŋt, a word in the Mansi languages of western Siberia meaning "earth horn", in reference to mammoth tusks.[4] Mammoths appear in the folkore of the indigenous people of Siberia, who were impressed by the great size of their remains. In the mythology of the Evenk people, mammoths were responsible for the creation of the world, digging up the land from the ocean floor with their tusks. The Selkup believed that mammoths lived underground and guarded the underworld, while the Nenets and the Mansi (the latter of whom, along with the Khanty, conceived mammoths as giant birds) believed that mammoths were responsible for the creation of mountains and lakes, while the Yakuts regarded mammoths as water spirits.[5]
The word mammoth was first used in Europe during the early 17th century, when referring to maimanto tusks discovered in Siberia,[6] as recorded in the 1618 edition of the Dictionariolum Russico-Anglicum.[7] The earliest scientific research paper on mammoths was by Vasily Tatishchev in 1725.[5] John Bell, who was on the Ob River in 1722, said that mammoth tusks were well known in the area. They were called "mammon's horn" and were often found in washed-out river banks. Bell bought one and presented it to Hans Sloan who pronounced it an elephant's tooth.[8]
In 1796, French biologist Georges Cuvier was the first to identify woolly mammoth remains not as modern elephants transported to the Arctic, but as an entirely new species. He argued this species had gone extinct and no longer existed, a concept that was not widely accepted at the time.[9][10] Following Cuvier's identification, German naturalist Johann Friedrich Blumenbach gave the woolly mammoth its scientific name, Elephas primigenius, in 1799, placing it in the same genus as the Asian elephant (Elephas maximus). This name is Latin for "the first-born elephant". Cuvier coined the name Elephas mammonteus a few months later, but the former name was subsequently used, including by Cuvier.[11] The genus name Mammuthus was coined by British anatomist Joshua Brookes in 1828, as part of a survey of his museum collection.[12]
Thomas Jefferson, who famously had a keen interest in paleontology, is partially responsible for transforming the word mammoth from a noun describing the prehistoric elephant to an adjective describing anything of surprisingly large size. The first recorded use of the word as an adjective was in a description of a large wheel of cheese (the "Cheshire Mammoth Cheese") given to Jefferson in 1802.[13]
Evolution
The earliest known proboscideans, the clade that contains the elephants, arose about 55 million years ago on the landmass of Afro-Arabia. The closest relatives of the Proboscidea are the sirenians and the hyraxes. The family Elephantidae arose by million years ago in Africa, and includes the living elephants and the mammoths. Among many now extinct clades, the mastodon is only a distant relative of the mammoths, and part of the separate Mammutidae family, which diverged 25 million years before the mammoths evolved.[14]
Following the publication of the woolly mammoths mitochondrial genome sequence in 1997, it has since become widely accepted that mammoths and Asian elephants share a closer relationship to each other than either do to African elephants.[15][16]
The following cladogram shows the placement of the genus Mammuthus among other proboscideans, based on hyoid characteristics and genetics:[17][16]
| ||||||||||||||||||||||||||||||||||||||||
It is possible to reconstruct the evolutionary history of the genus through morphological studies. Mammoth species can be identified from the number of enamel ridges/lamellae on their molars; the primitive species had few ridges, and the amount increased gradually as new species evolved and replaced the former ones. At the same time, the crowns of the teeth became longer, and the skulls became higher from top to bottom and shorter from the back to the front over time to accommodate this.[18]
The earliest mammoths, assigned to the species Mammuthus subplanifrons, are known from southern and eastern Africa, with the earliest records dating to the Late Miocene, around 6.2–5.3 million years ago.[3] By the Late Pliocene, mammoths had become confined to the northern portions of the African continent with remains from this time assigned to Mammuthus africanavus.[19] During the Late Pliocene, by 3.2 million years ago, mammoths dispersed into Eurasia via the Sinai Peninsula. The earliest mammoths in Eurasia are assigned to the species Mammuthus rumanus.[20] The youngest remains of mammoths in Africa are from Aïn Boucherit, Algeria dating to the Early Pleistocene, around 2.3–2 million years ago (with a possible later record from Aïn Hanech, Algeria, dating to 1.95–1.78 million years ago).[19]
Mammuthus rumanus is thought to be the ancestor of Mammuthus meridionalis, which first appeared at the beginning of the Pleistocene, around 2.6 million years ago.[21] Mammuthus meridionalis subsequently gave rise to Mammuthus trogontherii (the steppe mammoth) in Eastern Asia around 1.7 million years ago. Around 1.5–1.3 million years ago, M. trogontherii crossed the Bering Land Bridge into North America, becoming ancestral to Mammuthus columbi (the Columbian mammoth).[22] At the end of the Early Pleistocene Mammuthus trogontherii migrated into Europe, replacing M. meridionalis around 1–0.8 million years ago.[21] Mammuthus primigenius (the woolly mammoth) had evolved from M. trogontherii in Siberia by around 600,000–500,000 years ago, replacing M. trogontherii in Europe by around 200,000 years ago, and migrated into North America during the Late Pleistocene.[23]
A number of dwarf mammoth species, with small body sizes, evolved on islands as a result of insular dwarfism. These include Mammuthus lamarmorai on Sardinia (late Middle-Late Pleistocene),[24] Mammuthus exilis on the Channel Islands of California (Late Pleistocene),[25] and Mammuthus creticus on Crete (Early Pleistocene).[26]
Description
Like living elephants, mammoths typically had large body sizes. The largest known species like Mammuthus meridionalis and Mammuthus trogontherii (the steppe mammoth) were considerably larger than modern elephants, with mature adult males having an average height of approximately 3.8–4.2 m (12.5–13.8 ft) at the shoulder and weights of 9.6–12.7 tonnes (21,000–28,000 lb), while exceptionally large males may have reached 4.5 m (14.8 ft) at the shoulder and 14.3 tonnes (31,526.1 lb) in weight.[27] However, woolly mammoths were considerably smaller, only about as large as modern African bush elephants with males around 2.80–3.15 m (9 ft 2.2 in – 10 ft 4.0 in) high at the shoulder, and 4.5–6 tonnes (9,900–13,200 lb) in weight on average,[28] with the largest recorded individuals being around 3.5 m (11.5 ft) tall and 8.2 tonnes (18,077.9 lb) in weight.[27] The insular dwarf mammoth species were considerably smaller, with the smallest species M. creticus estimated to have a shoulder height of only around 1 metre (3.3 ft) and a weight of about 180 kilograms (400 lb), making it one of the smallest elephantids known.[27]
Gallery of mammoth skeletons

.png)
.png)

The number of lamellae (ridge-like structures) on the molars, particularly on the third molars, substantially increased over the course of mammoth evolution. The earliest Eurasian species M. rumanus have around 8-10 lamellae on the third molars,[29] while Late Pleistocene woolly mammoths have 20-28 lamellae on the third molars. These changes also corresponded with reduced enamel thickness and increasing tooth height (hypsodonty).[23] These changes are thought to be adaptations to increasing abrasion resulting from the shift in the diet of mammoths from a browsing based diet in M. rumanus, towards a grazing diet in later species.[30][31]
Molars

Cross section through elephantid molars, showing their internal structure
.jpg)
Molar of Mammuthus meridionalis
.jpg)
Molar of a woolly mammoth (Mammuthus primigenius)
Both sexes bore tusks. A first, small set appeared at about the age of six months, and these were replaced at about 18 months by the permanent set. Growth of the permanent set was at a rate of about 2.5 to 15.2 cm (1 to 6 in) per year.[32] The tusks display a strong spiral twisting.[33] Mammoth tusks are among the largest known among proboscideans with some specimens over 4 m (13.1 ft) in length and likely 200 kg (440.9 lb) in weight with some historical reports suggesting tusks of Columbian mammoths could reach lengths of around 5 m (16.4 ft) substantially surpassing the largest known modern elephant tusks.[34]
The heads of mammoths were prominently domed.[35] The first several thoracic vertebrae of mammoths typically had long neural spines.[36] The back was typically sloping, with the body being wider than that of African elephants. The tails of mammoths were relatively short compared to living elephants.[35]

While early mammoth species like M. meridionalis were probably relatively hairless, similar to modern elephants,[37] M. primigenius and likely M. trogontherii had a substantial coat of fur, among other physiological adaptations for living in cold environments. Genetic sequencing of M. trogontherii-like mammoths, over 1 million years old from Siberia suggests that they had already developed many of the genetic changes found in woolly mammoths responsible for tolerance of cold conditions.[38] Scientists discovered and studied the remains of a mammoth calf, and found that fat greatly influenced its form, and enabled it to store large amounts of nutrients necessary for survival in temperatures as low as −50 °C (−58 °F).[39] The fat also allowed the mammoths to increase their muscle mass, allowing the mammoths to fight against enemies and live longer.[40] Woolly mammoths evolved a suite of adaptations for arctic life, including morphological traits such as small ears and tails to minimize heat loss, a thick layer of subcutaneous fat, and numerous sebaceous glands for insulation, as well as a large brown-fat hump like deposit behind the neck that may have functioned as a heat source and fat reservoir during winter.[41]
Based on studies of their close relatives, the modern elephants, mammoths probably had a gestation period of 22 months, resulting in a single calf being born. Their social structure was probably the same as that of African and Asian elephants, with females living in herds headed by a matriarch, whilst bulls lived solitary lives or formed loose groups after sexual maturity,[42] with adult males experiencing periods of musth.[43]
Diet
The earliest mammoth species like M. subplanifrons and M. rumanus were mixed feeders (both browsing and grazing) to browsers. Over the course of mammoth evolution in Eurasia, their diet shifted towards mixed feeding-grazing in M. trogontherii, culminating in the woolly mammoth, which was largely a grazer, with stomach contents of woolly mammoths suggesting that they largely fed on grass and forbs. M. columbi is thought to have been a mixed feeder.[31]
Relationship with early humans

Evidence that humans interacted with mammoths extends back to around 1.8 million years ago, with a number of bones of Mammuthus meridionalis from the Dmanisi site in Georgia having marks suggested to the result of butchery by archaic humans, likely as a result of scavenging.[44] During the Last Glacial Period, modern humans hunted woolly mammoths,[45] used their remains to create art and tools,[46][45] and depicted them in works of art.[46] Remains of Columbian mammoths at a number of sites suggest that they were hunted by Paleoindians, the first humans to inhabit the Americas.[47] A possible bone engraving of a Columbian mammoth made by Paleoindians is known from Vero Beach, Florida.[48]
Extinction
Following the end of the Last Glacial Maximum, the range of the woolly mammoth began to contract, disappearing from most of Europe by 14,000 years ago.[49] By the Younger Dryas (around 12,900-11,700 years Before Present), woolly mammoths were confined to the northernmost regions of Siberia. This contraction is suggested to have been caused by the warming induced expansion of unfavourable wet tundra and forest environments at the expense of the preferred dry open mammoth steppe, with the possible additional pressure of human hunting. An additional issue under serious consideration is the impact of several asteroid or comet fragments impacting 12,800 years ago, evidence of which has been found in recent years, which resulted in forest fires and large dust emissions causing a sudden cooling of the climate (the Younger Dryas impact hypothesis) lasting about 1,400 years[50][51]. The last woolly mammoths in mainland Siberia became extinct around 10,000 years ago, during the early Holocene.[52] The final extinction of mainland woolly mammoths may have been driven by human hunting.[49] Relict populations survived on Saint Paul island in the Bering Strait until around 5,600 years ago, with their extinction likely due to the degradation of freshwater sources,[53] and on Wrangel Island off the coast of Northeast Siberia until around 4,000 years ago.[52] The last reliable dates of the Columbian mammoth date to around 12,500 years ago.[54] Columbian mammoths became extinct as part of the Late Pleistocene megafauna extinctions of most large mammals across the Americas approximately simultaneously at the end of the Late Pleistocene.[55] Hunting of Columbian mammoths by Paleoindians may have been a contributory factor in their extinction.[47] The timing of the extinction of the dwarf Sardinian mammoth Mammuthus lamarmorai is difficult to constrain precisely, though the youngest specimen likely dates to sometime around 57–29,000 years ago.[56] The youngest records of the pygmy mammoth (Mammuthus exillis) date to around 13,000 years ago, coinciding with the reducing of the area of the Californian Channel Islands as a result of rising sea level, the earliest known humans in the Channel Islands, and climatic change resulting in the decline of the previously dominant conifer forest ecosystems and expansion of scrub and grassland.[57]
See also
- Genesis 2.0, a documentary
- Ivory trade
- La Brea tar pits
- List of mammoths
- The Mammoth Site in Hot Springs, South Dakota
- Niederweningen Mammoth Museum
- Pleistocene Park
- Waco Mammoth National Monument
References
- ^ Garutt, W.E.; Gentry, Anthea; Lister, A.M. (1990). "Case 2726: Mammuthus Brookes 1828 (Mammalia Proboscidea) proposed conservation and Elephas primigenius Blumenbach, 1799 (currently Mammuthus primigenius) proposed designation as the type species of Mammuthus, and designation of a neotype". Bulletin of Zoological Nomenclature. 47 (1): 38–44. doi:10.5962/bhl.part.2651.
- ^ "Opinion 1661: Mammuthus Brookes, 1828 (Mammalia, Proboscidea): conserved, and Elephas primigenius Blumenbach, 1799 designated as the type species". Bulletin of Zoological Nomenclature. 48 (3): 279–280. 1991.
- ^ a b Sanders, William J. (2023-07-07). Evolution and Fossil Record of African Proboscidea (1 ed.). Boca Raton: CRC Press. pp. 155, 208–212. doi:10.1201/b20016. ISBN 978-1-315-11891-8. S2CID 259625811.
- ^ “mammoth”, in The American Heritage Dictionary of the English Language, 5th edition, Boston, Mass.: Houghton Mifflin Harcourt, 2016, →ISBN.
- ^ a b Serikov, Iu.B.; Serikova, A.Iu. (April 2005). "The Mammoth in the Myths, Ethnography, and Archeology of Northern Eurasia". Anthropology & Archeology of Eurasia. 43 (4): 8–18. doi:10.1080/10611959.2005.11029015. ISSN 1061-1959.
- ^ Lister, A.; Bahn, P. (2007). Mammoths – Giants of the Ice Age (3rd ed.). London: Frances Lincoln. p. 49. ISBN 978-0-520-26160-0.
- ^ "Mammoth" Oxford English Dictionary 2000
- ^ John Bell, Travels from St Petersburg in Russia to diverse parts of Asia, Edinburgh, 1806, pages 383-386
- ^ Switek, B. (2010). Written in Stone: Evolution, the Fossil Record, and Our Place in Nature. Bellevue Literary Press. pp. 174–180. ISBN 978-1-934137-29-1.
- ^ Cuvier, G. (1796). "Mémoire sur les épèces d'elephans tant vivantes que fossils, lu à la séance publique de l'Institut National le 15 germinal, an IV". Magasin Encyclopédique, 2e Anée (in French): 440–445.
- ^ Reich, M.; Gehler, A.; Mohl, D.; van der Plicht, H.; Lister, A. M. (2007). "The rediscovery of type material of Mammuthus primigenius (Mammalia: Proboscidea)". International Mammoth Conference IV (Poster): 295.
- ^ BROOKES, J., 1828. A catalogue of the anatomical and zoological museum of Jeshua Brookes, Esq., F.R.S. etc. Part 1. R.Taylor, London. 76 pp.
- ^ Simpson, J. (2009). "Word Stories: Mammoth." Oxford English Dictionary Online, Oxford University Press. Accessed 05-JUN-2009.
- ^ Lister, A.; Bahn, P. (2007). Mammoths – Giants of the Ice Age (3rd ed.). London: Frances Lincoln. ISBN 978-0-520-26160-0.
- ^ Ozawa, Tomowo; Hayashi, Seiji; Mikhelson, Victor M. (April 1997). "Phylogenetic Position of Mammoth and Steller's Sea Cow Within Tethytheria Demonstrated by Mitochondrial DNA Sequences". Journal of Molecular Evolution. 44 (4): 406–413. doi:10.1007/PL00006160. ISSN 0022-2844.
- ^ a b Palkopoulou, Eleftheria; Lipson, Mark; Mallick, Swapan; Nielsen, Svend; Rohland, Nadin; Baleka, Sina; Karpinski, Emil; Ivancevic, Atma M.; To, Thu-Hien; Kortschak, R. Daniel; Raison, Joy M. (2018-03-13). "A comprehensive genomic history of extinct and living elephants". Proceedings of the National Academy of Sciences. 115 (11): E2566–E2574. Bibcode:2018PNAS..115E2566P. doi:10.1073/pnas.1720554115. ISSN 0027-8424. PMC 5856550. PMID 29483247.
- ^ Shoshani, J.; Ferretti, M. P.; Lister, A. M.; Agenbroad, L. D.; Saegusa, H.; Mol, D.; Takahashi, K. (2007). "Relationships within the Elephantinae using hyoid characters". Quaternary International. 169–170: 174–185. Bibcode:2007QuInt.169..174S. doi:10.1016/j.quaint.2007.02.003.
- ^ Lister, A. M.; Sher, A. V.; Van Essen, H.; Wei, G. (2005). "The pattern and process of mammoth evolution in Eurasia" (PDF). Quaternary International. 126–128: 49–64. Bibcode:2005QuInt.126...49L. doi:10.1016/j.quaint.2004.04.014.
- ^ a b Sanders, William J. (2023-07-07). Evolution and Fossil Record of African Proboscidea (1 ed.). Boca Raton: CRC Press. pp. 245, 252, 263–266. doi:10.1201/b20016. ISBN 978-1-315-11891-8. S2CID 259625811.
- ^ Iannucci, Alessio; Sardella, Raffaele (2023-02-28). "What Does the "Elephant-Equus" Event Mean Today? Reflections on Mammal Dispersal Events around the Pliocene-Pleistocene Boundary and the Flexible Ambiguity of Biochronology". Quaternary. 6 (1): 16. doi:10.3390/quat6010016. hdl:11573/1680082. ISSN 2571-550X.
- ^ a b Lister, Adrian M.; Sher, Andrei V.; van Essen, Hans; Wei, Guangbiao (January 2005). "The pattern and process of mammoth evolution in Eurasia" (PDF). Quaternary International. 126–128: 49–64. Bibcode:2005QuInt.126...49L. doi:10.1016/j.quaint.2004.04.014. ISSN 1040-6182.
- ^ Lister, A. M.; Sher, A. V. (2015-11-13). "Evolution and dispersal of mammoths across the Northern Hemisphere". Science. 350 (6262): 805–809. Bibcode:2015Sci...350..805L. doi:10.1126/science.aac5660. ISSN 0036-8075. PMID 26564853. S2CID 206639522.
- ^ a b Lister, Adrian M. (October 2022). "Mammoth evolution in the late Middle Pleistocene: The Mammuthus trogontherii-primigenius transition in Europe". Quaternary Science Reviews. 294: 107693. Bibcode:2022QSRv..29407693L. doi:10.1016/j.quascirev.2022.107693. S2CID 252264887.
- ^ Palombo, Maria Rita; Zedda, Marco; Melis, Rita Teresa (November 2017). "A new elephant fossil from the late Pleistocene of Alghero: The puzzling question of Sardinian dwarf elephants". Comptes Rendus Palevol. 16 (8): 841–849. Bibcode:2017CRPal..16..841P. doi:10.1016/j.crpv.2017.05.007.
- ^ Agenbroad, L. D. (2010). ". Mammuthus exilis from the California Channel Islands: Height, Mass and Geologic Age" (PDF). Proceedings of the 7th California Islands Symposium. p. 17. Retrieved 13 June 2012.
- ^ Herridge, V. L.; Lister, A. M. (2012). "Extreme insular dwarfism evolved in a mammoth". Proceedings of the Royal Society B: Biological Sciences. 279 (1741): 3193–3300. doi:10.1098/rspb.2012.0671. PMC 3385739. PMID 22572206.
- ^ a b c Larramendi, A. (2016). "Shoulder height, body mass and shape of proboscideans" (PDF). Acta Palaeontologica Polonica. 61. doi:10.4202/app.00136.2014.
- ^ Larramendi, Asier; Palombo, Maria Rita; Marano, Federica (2017). "Reconstructing the life appearance of a Pleistocene giant: size, shape, sexual dimorphism and ontogeny of Palaeoloxodon antiquus (Proboscidea: Elephantidae) from Neumark-Nord 1 (Germany)" (PDF). Bollettino della Società Paleontologica Italiana (3): 299–317. doi:10.4435/BSPI.2017.29. ISSN 0375-7633. Archived from the original (PDF) on 2023-09-30.
- ^ Markov, Georgi N. (October 2012). "Mammuthus rumanus, early mammoths, and migration out of Africa: Some interrelated problems". Quaternary International. 276–277: 23–26. Bibcode:2012QuInt.276...23M. doi:10.1016/j.quaint.2011.05.041.
- ^ Lister, Adrian M.; Sher, Andrei V. (2001-11-02). "The Origin and Evolution of the Woolly Mammoth". Science. 294 (5544): 1094–1097. Bibcode:2001Sci...294.1094L. doi:10.1126/science.1056370. ISSN 0036-8075. PMID 11691991. S2CID 10662205.
- ^ a b Rivals, Florent; Semprebon, Gina M.; Lister, Adrian M. (September 2019). "Feeding traits and dietary variation in Pleistocene proboscideans: A tooth microwear review". Quaternary Science Reviews. 219: 145–153. Bibcode:2019QSRv..219..145R. doi:10.1016/j.quascirev.2019.06.027. S2CID 200073388.
- ^ Agenbroad, Larry; Nelson, Lisa (2002). Mammoths. Minneapolis: Lerner. p. 34. ISBN 978-0-8225-2862-3.
- ^ Rabinovich, Rivka; Lister, Adrian M. (July 2017). "The earliest elephants out of Africa: Taxonomy and taphonomy of proboscidean remains from Bethlehem". Quaternary International. 445: 23–42. Bibcode:2017QuInt.445...23R. doi:10.1016/j.quaint.2016.07.010.
- ^ Larramendi, Asier (2023-12-10). "Estimating tusk masses in proboscideans: a comprehensive analysis and predictive model". Historical Biology: 1–14. doi:10.1080/08912963.2023.2286272. ISSN 0891-2963. S2CID 266182491.
- ^ a b Larramendi, Asier; Palombo, Maria Rita; Marano, Federica (2017). "Reconstructing the life appearance of a Pleistocene giant: size, shape, sexual dimorphism and ontogeny of Palaeoloxodon antiquus (Proboscidea: Elephantidae) from Neumark-Nord 1 (Germany)" (PDF). Bollettino della Società Paleontologica Italiana (3): 299–317. doi:10.4435/BSPI.2017.29. ISSN 0375-7633. Archived from the original (PDF) on 2023-09-30.
- ^ Larramendi, Asier (2014-02-16). "Skeleton of a Late Pleistocene steppe mammoth (Mammuthus trogontherii) from Zhalainuoer, Inner Mongolian Autonomous Region, China". Paläontologische Zeitschrift. 89 (2): 229–250. doi:10.1007/s12542-014-0222-8. ISSN 0031-0220.
- ^ Lister, Adrian; Bahn, Paul (2007). Mammoths: giants of the ice age. Frances Lincoln LTD. pp. 25–26. ISBN 978-0-7112-2801-6.
- ^ van der Valk, Tom; Pečnerová, Patrícia; Díez-del-Molino, David; Bergström, Anders; Oppenheimer, Jonas; Hartmann, Stefanie; Xenikoudakis, Georgios; Thomas, Jessica A.; Dehasque, Marianne; Sağlıcan, Ekin; Fidan, Fatma Rabia (17 February 2021). "Million-year-old DNA sheds light on the genomic history of mammoths". Nature. 591 (7849): 265–269. Bibcode:2021Natur.591..265V. doi:10.1038/s41586-021-03224-9. ISSN 1476-4687. PMC 7116897. PMID 33597750.
- ^ Peter D. Moore (2008). Tundra. Facts On File. p. 198. ISBN 978-0-8160-5933-1.
- ^ Maschenko, E. N.; Boeskorov, G. G.; Baranov, V. A. (2013). "Morphology of a mammoth calf (Mammuthus primigenius) from Ol'chan (Oimiakon, Yakutia)". Paleontological Journal. 47 (4): 425–438. Bibcode:2013PalJ...47..425M. doi:10.1134/S0031030113040096. S2CID 84317574.
- ^ Lynch, Vincent (2 July 2015). "Elephantid Genomes Reveal the Molecular Bases of Woolly Mammoth Adaptations to the Arctic". Cell Reports. 12 (2): P217-228. doi:10.1016/j.celrep.2015.06.027. hdl:10220/38768. PMID 26146078.
- ^ "Columbian Mammoth & Channel Island Mammoth". San Diego Zoo. Archived from the original on 2011-07-27. Retrieved 2010-06-15.
- ^ Cherney, Michael D.; Fisher, Daniel C.; Auchus, Richard J.; Rountrey, Adam N.; Selcer, Perrin; Shirley, Ethan A.; Beld, Scott G.; Buigues, Bernard; Mol, Dick; Boeskorov, Gennady G.; Vartanyan, Sergey L.; Tikhonov, Alexei N. (2023-05-18). "Testosterone histories from tusks reveal woolly mammoth musth episodes". Nature. 617 (7961): 533–539. Bibcode:2023Natur.617..533C. doi:10.1038/s41586-023-06020-9. ISSN 0028-0836. PMID 37138076. S2CID 258485513.
- ^ Tappen, Martha; Bukhsianidze, Maia; Ferring, Reid; Coil, Reed; Lordkipanidze, David (October 2022). "Life and death at Dmanisi, Georgia: Taphonomic signals from the fossil mammals". Journal of Human Evolution. 171: 103249. Bibcode:2022JHumE.17103249T. doi:10.1016/j.jhevol.2022.103249. PMID 36116366.
- ^ a b Wojtal, Piotr; Wilczyński, Jarosław (August 2015). "Hunters of the giants: Woolly mammoth hunting during the Gravettian in Central Europe". Quaternary International. 379: 71–81. Bibcode:2015QuInt.379...71W. doi:10.1016/j.quaint.2015.05.040.
- ^ a b Braun, Ingmar M.; Palombo, Maria Rita (October 2012). "Mammuthus primigenius in the cave and portable art: An overview with a short account on the elephant fossil record in Southern Europe during the last glacial". Quaternary International. 276–277: 61–76. Bibcode:2012QuInt.276...61B. doi:10.1016/j.quaint.2012.07.010.
- ^ a b Haynes, Gary (2022-07-03). "Sites in the Americas with Possible or Probable Evidence for the Butchering of Proboscideans". PaleoAmerica. 8 (3): 187–214. doi:10.1080/20555563.2022.2057834. ISSN 2055-5563. S2CID 251042359.
- ^ Purdy, Barbara A.; Jones, Kevin S.; Mecholsky, John J.; Bourne, Gerald; Hulbert, Richard C.; MacFadden, Bruce J.; Church, Krista L.; Warren, Michael W.; Jorstad, Thomas F.; Stanford, Dennis J.; Wachowiak, Melvin J.; Speakman, Robert J. (November 2011). "Earliest art in the Americas: incised image of a proboscidean on a mineralized extinct animal bone from Vero Beach, Florida". Journal of Archaeological Science. 38 (11): 2908–2913. Bibcode:2011JArSc..38.2908P. doi:10.1016/j.jas.2011.05.022.
- ^ a b Fordham, Damien A.; Brown, Stuart C.; Akçakaya, H. Reşit; Brook, Barry W.; Haythorne, Sean; Manica, Andrea; Shoemaker, Kevin T.; Austin, Jeremy J.; Blonder, Benjamin; Pilowsky, July A.; Rahbek, Carsten; Nogues-Bravo, David (January 2022). Coulson, Tim (ed.). "Process-explicit models reveal pathway to extinction for woolly mammoth using pattern-oriented validation". Ecology Letters. 25 (1): 125–137. Bibcode:2022EcolL..25..125F. doi:10.1111/ele.13911. hdl:11343/299174. ISSN 1461-023X. PMID 34738712.
- ^ Ward, C.J.G. (22 October 2019). "UofSC archaeologist finds evidence of extinction theory". University of South Carolina. Retrieved 21 February 2024.
- ^ Holliday, V.T.; Daulton, T.L; Bartlein, P.J.; Boslough, M.B.; Breslawski, R.P.; Fisher, A.E.; Jorgeson, I.A.; Scott, A.C.; Koeberl, C.; Marlon, J.R.; Severinghaus, J.; Petaev, M.I.; Claeys, P. (December 2023). "Comprehensive refutation of the Younger Dryas Impact Hypothesis (YDIH)". Earth-Science Reviews. Retrieved 21 February 2024.
- ^ a b Dehasque, Marianne; Pečnerová, Patrícia; Muller, Héloïse; Tikhonov, Alexei; Nikolskiy, Pavel; Tsigankova, Valeriya I.; Danilov, Gleb K.; Díez-del-Molino, David; Vartanyan, Sergey; Dalén, Love; Lister, Adrian M. (May 2021). "Combining Bayesian age models and genetics to investigate population dynamics and extinction of the last mammoths in northern Siberia". Quaternary Science Reviews. 259: 106913. Bibcode:2021QSRv..25906913D. doi:10.1016/j.quascirev.2021.106913.
- ^ Graham, Russell W.; Belmecheri, Soumaya; Choy, Kyungcheol; Culleton, Brendan J.; Davies, Lauren J.; Froese, Duane; Heintzman, Peter D.; Hritz, Carrie; Kapp, Joshua D.; Newsom, Lee A.; Rawcliffe, Ruth; Saulnier-Talbot, Émilie; Shapiro, Beth; Wang, Yue; Williams, John W. (2016-08-16). "Timing and causes of mid-Holocene mammoth extinction on St. Paul Island, Alaska". Proceedings of the National Academy of Sciences. 113 (33): 9310–9314. Bibcode:2016PNAS..113.9310G. doi:10.1073/pnas.1604903113. ISSN 0027-8424. PMC 4995940. PMID 27482085.
- ^ Stuart, Anthony J. "North America: Mastodon, Ground Sloths, and Sabertooth Cats". Vanished Giants: The Lost World of the Ice Age. Chicago, IL: University of Chicago Press. p. 97.
- ^ Faith, J. Tyler; Surovell, Todd A. (2009-12-08). "Synchronous extinction of North America's Pleistocene mammals". Proceedings of the National Academy of Sciences. 106 (49): 20641–20645. Bibcode:2009PNAS..10620641F. doi:10.1073/pnas.0908153106. ISSN 0027-8424. PMC 2791611. PMID 19934040.
- ^ Palombo, Maria Rita; Zedda, Marco; Zoboli, Daniel (March 2024). "The Sardinian Mammoth's Evolutionary History: Lights and Shadows". Quaternary. 7 (1): 10. doi:10.3390/quat7010010. ISSN 2571-550X.
- ^ Semprebon, Gina M.; Rivals, Florent; Fahlke, Julia M.; Sanders, William J.; Lister, Adrian M.; Göhlich, Ursula B. (June 2016). "Dietary reconstruction of pygmy mammoths from Santa Rosa Island of California". Quaternary International. 406: 123–136. doi:10.1016/j.quaint.2015.10.120.
Further reading
Wikimedia Commons has media related to Mammuthus.
Wikispecies has information related to Mammuthus.
Look up mammoth in Wiktionary, the free dictionary.
- Bahn, Paul G.; Lister, Adrian (1994). Mammoths. New York: Macmillan USA. ISBN 978-0-02-572985-8.
- Capelli, C.; MacPhee, R. D. E.; Roca, A. L.; Brisighelli, F.; Georgiadis, N.; O'Brien, S. J.; Greenwood, A. D. (2006). "A nuclear DNA phylogeny of the woolly mammoth (Mammuthus primigenius)". Molecular Phylogenetics and Evolution. 40 (2): 620–627. Bibcode:2006MolPE..40..620C. doi:10.1016/j.ympev.2006.03.015. PMID 16631387.
- Conniff, R. (2010). "Mammoths and Mastodons: All American Monsters". Smithsonian Magazine. Retrieved 2012-03-07.
- "Mammoth genome cracked: key to cloning". COSMOS magazine. 2008. Archived from the original on 2012-03-22. Retrieved 2012-03-07.
- "National Park Service Findings 'Good News' For Waco Mammoth Site". Baylor University. 2007-03-27. Retrieved 2012-03-07.
- Hayes, J. (2006). "Back from the dead". COSMOS magazine. Archived from the original on 2012-03-22. Retrieved 2012-03-07.
- Haynes, G. (1991). Mammoths, mastodons, and elephants. Biology, behavior, and the fossil record. Cambridge: Cambridge University Press. ISBN 0-521-38435-4.
- Keddie, G. "The Mammoth Story" (PDF). Royal BC Museum. Archived from the original (PDF) on 2011-12-25. Retrieved 2012-03-07.
- Levy, S. (2006). "Clashing with Titans". BioScience. 56 (4): 292. doi:10.1641/0006-3568(2006)56[292:CWT]2.0.CO;2.
- Martin, Paul (2005). Twilight of the mammoths: ice age extinctions and the rewilding of America. Berkeley: University of California Press. ISBN 978-0-520-23141-2.
- Mercer, Henry Chapman (2010) [1885]. The Lenape Stone: Or The Indian And The Mammoth (1885). Kessinger Publishing, LLC. ISBN 978-1-161-69753-7.
- Rodgers, J. (2006). "Mammoth skeleton found in Siberia". BBC News. Retrieved 2012-03-07.
- Stone, Richard G. (2003). Mammoth: The Resurrection of an Ice Age Giant. Fourth Estate Ltd. ISBN 978-1-84115-518-0.
| General |
| ||||||
|---|---|---|---|---|---|---|---|
| Human use |
| ||||||
| Culture and history |
| ||||||
| Related | |||||||
Genera of the order Proboscidea | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||
| 
 | |||||||||||||||||||||||||||||
