Though estradiol levels in males are much lower than in females, estradiol has important roles in males as well. Apart from humans and other mammals, estradiol is also found in most vertebrates and crustaceans, insects, fish, and other animalspecies.[8][9]
Estradiol is produced especially within the follicles of the ovaries, but also in other tissues including the testicles, the adrenal glands, fat, liver, the breasts, and the brain. Estradiol is produced in the body from cholesterol through a series of reactions and intermediates.[10] The major pathway involves the formation of androstenedione, which is then converted by aromatase into estrone and is subsequently converted into estradiol. Alternatively, androstenedione can be converted into testosterone, which can then be converted into estradiol. Upon menopause in females, production of estrogens by the ovaries stops and estradiol levels decrease to very low levels.
In the female, estradiol acts as a growth hormone for tissue of the reproductive organs, supporting the lining of the vagina, the cervical glands, the endometrium, and the lining of the fallopian tubes. It enhances growth of the myometrium. Estradiol appears necessary to maintain oocytes in the ovary. During the menstrual cycle, estradiol produced by the growing follicles triggers, via a positive feedback system, the hypothalamic-pituitary events that lead to the luteinizing hormone surge, inducing ovulation. In the luteal phase, estradiol, in conjunction with progesterone, prepares the endometrium for implantation. During pregnancy, estradiol increases due to placental production. The effect of estradiol, together with estrone and estriol, in pregnancy is less clear. They may promote uterine blood flow, myometrial growth, stimulate breast growth and at term, promote cervical softening and expression of myometrial oxytocin receptors.[citation needed] In baboons, blocking of estrogen production leads to pregnancy loss, suggesting estradiol has a role in the maintenance of pregnancy. Research is investigating the role of estrogens in the process of initiation of labor. Actions of estradiol are required before the exposure of progesterone in the luteal phase.[citation needed]
Male reproductive system
The effect of estradiol (and estrogens in general) upon male reproduction is complex. Estradiol is produced by action of aromatase mainly in the Leydig cells of the mammaliantestis, but also by some germ cells and the Sertoli cells of immature mammals.[14] It functions (in vitro) to prevent apoptosis of male sperm cells.[15]
While some studies in the early 1990s claimed a connection between globally declining sperm counts and estrogen exposure in the environment,[16] later studies found no such connection, nor evidence of a general decline in sperm counts.[17][18]
Suppression of estradiol production in a subpopulation of subfertile men may improve the semen analysis.[19]
Estradiol has a profound effect on bone. Individuals without it (or other estrogens) will become tall and eunuchoid, as epiphyseal closure is delayed or may not take place.[21]Bone density is also affected, resulting in early osteopenia and osteoporosis.[22] Low levels of estradiol may also predict fractures, with post-menopausal women having the highest incidence of bone fracture.[23] Women past menopause experience an accelerated loss of bone mass due to a relative estrogen deficiency.[24]
Hormone replacement therapy consisting of systemic treatment with estrogen alone or in combination with a progestogen, has well-documented and considerable beneficial effects on the skin of postmenopausal women.[25][26] These benefits include increased skin collagen content, skin thickness and elasticity, and skin hydration and surface lipids.[25][26] Topical estrogen has been found to have similar beneficial effects on the skin.[25] In addition, a study has found that topical 2% progesterone cream significantly increases skin elasticity and firmness and observably decreases wrinkles in peri- and postmenopausal women.[26] Skin hydration and surface lipids, on the other hand, did not significantly change with topical progesterone.[26] These findings suggest that progesterone, like estrogen, also has beneficial effects on the skin, and may be independently protective against skin aging.[26]
Estrogen is considered to play a significant role in women's mental health, with links suggested between the hormone level, mood and well-being. Sudden drops or fluctuations in, or long periods of sustained low levels of estrogen may be correlated with significant mood-lowering. Clinical recovery from depression postpartum, perimenopause, and postmenopause was shown to be effective after levels of estrogen were stabilized and/or restored.[29][30]
The volumes of sexually dimorphic brain structures in transgender women were found to change and approximate typical female brain structures when exposed to estrogen concomitantly with androgen deprivation over a period of months,[31] suggesting that estrogen and/or androgens have a significant part to play in sex differentiation of the brain, both prenatally and later in life.
There is also evidence the programming of adult male sexual behavior in many vertebrates is largely dependent on estradiol produced during prenatal life and early infancy.[32] It is not yet known whether this process plays a significant role in human sexual behavior, although evidence from other mammals tends to indicate a connection.[33]
Estrogen has been found to increase the secretion of oxytocin and to increase the expression of its receptor, the oxytocin receptor, in the brain.[34] In women, a single dose of estradiol has been found to be sufficient to increase circulating oxytocin concentrations.[35]
Gynecological cancers
Estradiol has been tied to the development and progression of cancers such as breast cancer, ovarian cancer and endometrial cancer. Estradiol affects target tissues mainly by interacting with two nuclear receptors called estrogen receptor α (ERα) and estrogen receptor β (ERβ).[36][37] One of the functions of these estrogen receptors is the modulation of gene expression. Once estradiol binds to the ERs, the receptor complexes then bind to specific DNA sequences, possibly causing damage to the DNA and an increase in cell division and DNA replication. Eukaryotic cells respond to damaged DNA by stimulating or impairing G1, S, or G2 phases of the cell cycle to initiate DNA repair. As a result, cellular transformation and cancer cell proliferation occurs.[38]
Cardiovascular system
Estrogen affects certain blood vessels. Improvement in arterial blood flow has been demonstrated in coronary arteries.[39] 17-beta-estradiol (E2) is considered the most potent estrogen found in humans. E2 influences vascular function, apoptosis, and damage during cardiac ischemia and reperfusion. E2 can protect the heart and individual cardiac myocytes from injuries related to ischemia. After a heart attack or long periods of hypertension, E2 inhibits the adverse effects of pathologic remodeling of the heart.[40]
Absolute and relative incidence of venous thromboembolism (VTE) during pregnancy and the postpartum period
Absolute incidence of first VTE per 10,000 person–years during pregnancy and the postpartum period
Swedish data A
Swedish data B
English data
Danish data
Time period
N
Rate (95% CI)
N
Rate (95% CI)
N
Rate (95% CI)
N
Rate (95% CI)
Outside pregnancy
1105
4.2 (4.0–4.4)
1015
3.8 (?)
1480
3.2 (3.0–3.3)
2895
3.6 (3.4–3.7)
Antepartum
995
20.5 (19.2–21.8)
690
14.2 (13.2–15.3)
156
9.9 (8.5–11.6)
491
10.7 (9.7–11.6)
Trimester 1
207
13.6 (11.8–15.5)
172
11.3 (9.7–13.1)
23
4.6 (3.1–7.0)
61
4.1 (3.2–5.2)
Trimester 2
275
17.4 (15.4–19.6)
178
11.2 (9.7–13.0)
30
5.8 (4.1–8.3)
75
5.7 (4.6–7.2)
Trimester 3
513
29.2 (26.8–31.9)
340
19.4 (17.4–21.6)
103
18.2 (15.0–22.1)
355
19.7 (17.7–21.9)
Around delivery
115
154.6 (128.8–185.6)
79
106.1 (85.1–132.3)
34
142.8 (102.0–199.8)
–
Postpartum
649
42.3 (39.2–45.7)
509
33.1 (30.4–36.1)
135
27.4 (23.1–32.4)
218
17.5 (15.3–20.0)
Early postpartum
584
75.4 (69.6–81.8)
460
59.3 (54.1–65.0)
177
46.8 (39.1–56.1)
199
30.4 (26.4–35.0)
Late postpartum
65
8.5 (7.0–10.9)
49
6.4 (4.9–8.5)
18
7.3 (4.6–11.6)
319
3.2 (1.9–5.0)
Incidence rate ratios (IRRs) of first VTE during pregnancy and the postpartum period
Swedish data A
Swedish data B
English data
Danish data
Time period
IRR* (95% CI)
IRR* (95% CI)
IRR (95% CI)†
IRR (95% CI)†
Outside pregnancy
Reference (i.e., 1.00)
Antepartum
5.08 (4.66–5.54)
3.80 (3.44–4.19)
3.10 (2.63–3.66)
2.95 (2.68–3.25)
Trimester 1
3.42 (2.95–3.98)
3.04 (2.58–3.56)
1.46 (0.96–2.20)
1.12 (0.86–1.45)
Trimester 2
4.31 (3.78–4.93)
3.01 (2.56–3.53)
1.82 (1.27–2.62)
1.58 (1.24–1.99)
Trimester 3
7.14 (6.43–7.94)
5.12 (4.53–5.80)
5.69 (4.66–6.95)
5.48 (4.89–6.12)
Around delivery
37.5 (30.9–44.45)
27.97 (22.24–35.17)
44.5 (31.68–62.54)
–
Postpartum
10.21 (9.27–11.25)
8.72 (7.83–9.70)
8.54 (7.16–10.19)
4.85 (4.21–5.57)
Early postpartum
19.27 (16.53–20.21)
15.62 (14.00–17.45)
14.61 (12.10–17.67)
8.44 (7.27–9.75)
Late postpartum
2.06 (1.60–2.64)
1.69 (1.26–2.25)
2.29 (1.44–3.65)
0.89 (0.53–1.39)
Notes: Swedish data A = Using any code for VTE regardless of confirmation. Swedish data B = Using only algorithm-confirmed VTE. Early postpartum = First 6 weeks after delivery. Late postpartum = More than 6 weeks after delivery. * = Adjusted for age and calendar year. † = Unadjusted ratio calculated based on the data provided. Source:[41]
Estradiol acts primarily as an agonist of the estrogen receptor (ER), a nuclearsteroid hormone receptor. There are two subtypes of the ER, ERα and ERβ, and estradiol potently binds to and activates both of these receptors. The result of ER activation is a modulation of gene transcription and expression in ER-expressing cells, which is the predominant mechanism by which estradiol mediates its biological effects in the body. Estradiol also acts as an agonist of membrane estrogen receptors (mERs), such as GPER (GPR30), a recently discovered non-nuclear receptor for estradiol, via which it can mediate a variety of rapid, non-genomic effects.[42] Unlike the case of the ER, GPER appears to be selective for estradiol, and shows very low affinities for other endogenous estrogens, such as estrone and estriol.[43] Additional mERs besides GPER include ER-X, ERx, and Gq-mER.[44][45]
ERα/ERβ are in inactive state trapped in multimolecular chaperone complexes organized around the heat shock protein 90 (HSP90), containing p23 protein, and immunophilin, and located in majority in cytoplasm and partially in nucleus. In the E2 classical pathway or estrogen classical pathway, estradiol enters the cytoplasm, where it interacts with ERs. Once bound E2, ERs dissociate from the molecular chaperone complexes and become competent to dimerize, migrate to nucleus, and to bind to specific DNA sequences (estrogen response element, ERE), allowing for gene transcription which can take place over hours and days.
Given by subcutaneous injection in mice, estradiol is about 10-fold more potent than estrone and about 100-fold more potent than estriol.[46][47][48] As such, estradiol is the main estrogen in the body, although the roles of estrone and estriol as estrogens are said not to be negligible.[48]
Notes: Values are mean ± SD or range. ER RBA = Relative binding affinity to estrogen receptors of rat uterinecytosol. Uterine weight = Percentage change in uterine wet weight of ovariectomized rats after 72 hours with continuous administration of 1 μg/hour via subcutaneously implantedosmotic pumps. LH levels = Luteinizing hormone levels relative to baseline of ovariectomized rats after 24 to 72 hours of continuous administration via subcutaneous implant. Footnotes:a = Synthetic (i.e., not endogenous). b = Atypical uterotrophic effect which plateaus within 48 hours (estradiol's uterotrophy continues linearly up to 72 hours). Sources: See template.
Estradiol, like other steroid hormones, is derived from cholesterol. After side chain cleavage and using the Δ5 or the Δ4- pathway, androstenedione is the key intermediary. A portion of the androstenedione is converted to testosterone, which in turn undergoes conversion to estradiol by aromatase. In an alternative pathway, androstenedione is aromatized to estrone, which is subsequently converted to estradiol via 17β-hydroxysteroid dehydrogenase (17β-HSD).[50]
During the reproductive years, most estradiol in women is produced by the granulosa cells of the ovaries by the aromatization of androstenedione (produced in the theca folliculi cells) to estrone, followed by conversion of estrone to estradiol by 17β-HSD. Smaller amounts of estradiol are also produced by the adrenal cortex, and, in men, by the testes.[medical citation needed]
Estradiol is not produced in the gonads only; in particular, fat cells produce active precursors to estradiol, and will continue to do so even after menopause.[51] Estradiol is also produced in the brain and in arterial walls.
In men, approximately 15 to 25% of circulating estradiol is produced in the testicles.[52][53] The rest is synthesized via peripheral aromatization of testosterone into estradiol and of androstenedione into estrone (which is then transformed into estradiol via peripheral 17β-HSD).[52][53] This peripheral aromatization occurs predominantly in adipose tissue, but also occurs in other tissues such as bone, liver, and the brain.[52] Approximately 40 to 50 μg of estradiol is produced per day in men.[52]
Distribution
In plasma, estradiol is largely bound to SHBG and albumin. Only about 2.21% (± 0.04%) of estradiol is free and biologically active. The percentage remains constant throughout the menstrual cycle.[54]
Estradiol levels across the menstrual cycle in 36 normally cycling, ovulatory women, based on 956 specimens.[61] The horizontal dashed lines are the mean integrated levels for each curve. The vertical dashed line in the center is mid-cycle.
Levels of estradiol in premenopausal women are highly variable throughout the menstrual cycle and reference ranges widely vary from source to source.[62] Estradiol levels are minimal and according to most laboratories range from 20 to 80 pg/mL during the early to mid follicular phase (or the first week of the menstrual cycle, also known as menses).[63][64] Levels of estradiol gradually increase during this time and through the mid to late follicular phase (or the second week of the menstrual cycle) until the pre-ovulatory phase.[62][63] At the time of pre-ovulation (a period of about 24 to 48 hours), estradiol levels briefly surge and reach their highest concentrations of any other time during the menstrual cycle.[62] Circulating levels are typically between 130 and 200 pg/mL at this time, but in some women may be as high as 300 to 400 pg/mL, and the upper limit of the reference range of some laboratories are even greater (for instance, 750 pg/mL).[62][63][65][66][67] Following ovulation (or mid-cycle) and during the latter half of the menstrual cycle or the luteal phase, estradiol levels plateau and fluctuate between around 100 and 150 pg/mL during the early and mid luteal phase, and at the time of the late luteal phase, or a few days before menstruation, reach a low of around 40 pg/mL.[62][64] The mean integrated levels of estradiol during a full menstrual cycle have variously been reported by different sources as 80, 120, and 150 pg/mL.[64][68][69] Although contradictory reports exist, one study found mean integrated estradiol levels of 150 pg/mL in younger women whereas mean integrated levels ranged from 50 to 120 pg/mL in older women.[69]
During the reproductive years of human females, levels of estradiol are somewhat higher than that of estrone, except during the early follicular phase of the menstrual cycle; thus, estradiol may be considered the predominant estrogen during human female reproductive years in terms of absolute serum levels and estrogenic activity.[citation needed] During pregnancy, estriol becomes the predominant circulating estrogen, and this is the only time at which estetrol occurs in the body, while during menopause, estrone predominates (both based on serum levels).[citation needed] The estradiol produced by male humans, from testosterone, is present at serum levels roughly comparable to those of postmenopausal women (14–55 versus <35 pg/mL, respectively).[citation needed] It has also been reported that if concentrations of estradiol in a 70-year-old man are compared to those of a 70-year-old woman, levels are approximately 2- to 4-fold higher in the man.[70]
In women, serum estradiol is measured in a clinical laboratory and reflects primarily the activity of the ovaries. The Estradiol blood test measures the amount of estradiol in the blood.[77] It is used to check the function of the ovaries, placenta, adrenal glands.[77] This can detect baseline estrogen in women with amenorrhea or menstrual dysfunction, and to detect the state of hypoestrogenicity and menopause. Furthermore, estrogen monitoring during fertility therapy assesses follicular growth and is useful in monitoring the treatment. Estrogen-producing tumors will demonstrate persistent high levels of estradiol and other estrogens. In precocious puberty, estradiol levels are inappropriately increased.
Ranges
Individual laboratory results should always be interpreted using the ranges provided by the laboratory that performed the test.
Reference ranges for the blood content of estradiol during the menstrual cycle
Reference ranges for the blood content of estradiol during the menstrual cycle - The ranges denoted By biological stage may be used in closely monitored menstrual cycles in regard to other markers of its biological progression, with the time scale being compressed or stretched to how much faster or slower, respectively, the cycle progresses compared to an average cycle. - The ranges denoted Inter-cycle variability are more appropriate to use in unmonitored cycles with only the beginning of menstruation known, but where the woman accurately knows her average cycle lengths and time of ovulation, and that they are somewhat averagely regular, with the time scale being compressed or stretched to how much a woman's average cycle length is shorter or longer, respectively, than the average of the population. - The ranges denoted Inter-woman variability are more appropriate to use when the average cycle lengths and time of ovulation are unknown, but only the beginning of menstruation is given.[81]
In the normal menstrual cycle, estradiol levels measure typically <50 pg/mL at menstruation, rise with follicular development (peak: 200 pg/mL), drop briefly at ovulation, and rise again during the luteal phase for a second peak. At the end of the luteal phase, estradiol levels drop to their menstrual levels unless there is a pregnancy.
During pregnancy, estrogen levels, including estradiol, rise steadily toward term. The source of these estrogens is the placenta, which aromatizes prohormones produced in the fetal adrenal gland.
Notes: "The concentration of a steroid in the circulation is determined by the rate at which it is secreted from glands, the rate of metabolism of precursor or prehormones into the steroid, and the rate at which it is extracted by tissues and metabolized. The secretion rate of a steroid refers to the total secretion of the compound from a gland per unit time. Secretion rates have been assessed by sampling the venous effluent from a gland over time and subtracting out the arterial and peripheral venous hormone concentration. The metabolic clearance rate of a steroid is defined as the volume of blood that has been completely cleared of the hormone per unit time. The production rate of a steroid hormone refers to entry into the blood of the compound from all possible sources, including secretion from glands and conversion of prohormones into the steroid of interest. At steady state, the amount of hormone entering the blood from all sources will be equal to the rate at which it is being cleared (metabolic clearance rate) multiplied by blood concentration (production rate = metabolic clearance rate × concentration). If there is little contribution of prohormone metabolism to the circulating pool of steroid, then the production rate will approximate the secretion rate." Sources: See template.
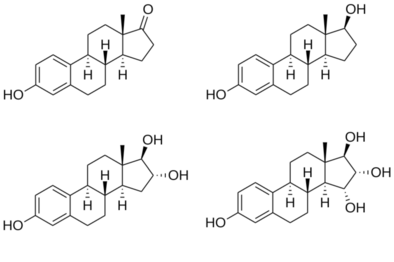
Note the hydroxyl (–OH) groups: estrone (E1) has one, estradiol (E2) has two, estriol (E3) has three, and estetrol (E4) has four.
Estradiol is an estranesteroid.[82] It is also known as 17β-estradiol (to distinguish it from 17α-estradiol) or as estra-1,3,5(10)-triene-3,17β-diol. It has two hydroxyl groups, one at the C3 position and the other at the 17β position, as well as three double bonds in the A ring. Due to its two hydroxyl groups, estradiol is often abbreviated as E2. The structurally related estrogens, estrone (E1), estriol (E3), and estetrol (E4) have one, three, and four hydroxyl groups, respectively.
Neuropsychopharmacology
In a randomized, double-blind, placebo-controlled study, estradiol was shown to have gender-specific effects on fairness sensitivity. Overall, when the division of a given amount of money was framed as either fair or unfair in a modified version of the ultimatum game, estradiol increased the acceptance rate of fair-framed proposals among men and decreased it among women. However, among the placebo-group "the mere belief of receiving estradiol treatment significantly increased the acceptance of unfair-framed offers in both sexes", indicating that so-called "environmental" factors played a role in organising the responses towards these presentations of the ultimatum game.[83]
Estradiol, the most potent of the three major estrogens, was the last of the three to be identified.[85][89] It was discovered by Schwenk and Hildebrant in 1933, who synthesized it via reduction of estrone.[85] Estradiol was subsequently isolated and purified from sow ovaries by Doisy in 1935, with its chemical structure determined simultaneously,[92] and was referred to variously as dihydrotheelin, dihydrofolliculin, dihydrofollicular hormone, and dihydroxyestrin.[85][93][94] In 1935, the name estradiol and the term estrogen were formally established by the Sex Hormone Committee of the Health Organization of the League of Nations; this followed the names estrone (which was initially called theelin, progynon, folliculin, and ketohydroxyestrin) and estriol (initially called theelol and trihydroxyestrin) having been established in 1932 at the first meeting of the International Conference on the Standardization of Sex Hormones in London.[89][95] Following its discovery, a partial synthesis of estradiol from cholesterol was developed by Inhoffen and Hohlweg in 1940, and a total synthesis was developed by Anner and Miescher in 1948.[85]
Society and culture
Etymology
The name estradiol derives from estra-, Gk.οἶστρος (oistros, literally meaning "verve or inspiration"),[96] which refers to the estranesteroidring system, and -diol, a chemical term and suffix indicating that the compound is a type of alcohol bearing two hydroxylgroups.
^Price TM, Blauer KL, Hansen M, Stanczyk F, Lobo R, Bates GW (March 1997). "Single-dose pharmacokinetics of sublingual versus oral administration of micronized 17 beta-estradiol". Obstetrics and Gynecology. 89 (3): 340–5. doi:10.1016/S0029-7844(96)00513-3. PMID9052581. S2CID71641652.
^Naunton M, Al Hadithy AF, Brouwers JR, Archer DF (2006). "Estradiol gel: review of the pharmacology, pharmacokinetics, efficacy, and safety in menopausal women". Menopause. 13 (3): 517–27. doi:10.1097/01.gme.0000191881.52175.8c. PMID16735950. S2CID42748448.
^Ryan KJ (August 1982). "Biochemistry of aromatase: significance to female reproductive physiology". Cancer Research. 42 (8 Suppl): 3342s–3344s. PMID7083198.
^Mechoulam R, Brueggemeier RW, Denlinger DL (September 1984). "Estrogens in insects". Cellular and Molecular Life Sciences. 40 (9): 942–944. doi:10.1007/BF01946450. S2CID31950471.
^Saldanha, Colin J., Luke Remage-Healey, and Barney A. Schlinger. "Synaptocrine signaling: steroid synthesis and action at the synapse." Endocrine reviews 32.4 (2011): 532–549.
^Sharpe RM, Skakkebaek NE (May 1993). "Are oestrogens involved in falling sperm counts and disorders of the male reproductive tract?". Lancet. 341 (8857): 1392–5. doi:10.1016/0140-6736(93)90953-E. PMID8098802. S2CID33135527.
^Handelsman DJ (2001). "Estrogens and falling sperm counts". Reproduction, Fertility, and Development. 13 (4): 317–24. doi:10.1071/rd00103. PMID11800170.
^ abcdefghHolzer G, Riegler E, Hönigsmann H, Farokhnia S, Schmidt JB, Schmidt B (September 2005). "Effects and side-effects of 2% progesterone cream on the skin of peri- and postmenopausal women: results from a double-blind, vehicle-controlled, randomized study". The British Journal of Dermatology. 153 (3): 626–34. doi:10.1111/j.1365-2133.2005.06685.x. PMID16120154. S2CID6077829.
^Behl C, Widmann M, Trapp T, Holsboer F (November 1995). "17-beta estradiol protects neurons from oxidative stress-induced cell death in vitro". Biochemical and Biophysical Research Communications. 216 (2): 473–82. doi:10.1006/bbrc.1995.2647. PMID7488136.
^Douma SL, Husband C, O'Donnell ME, Barwin BN, Woodend AK (2005). "Estrogen-related mood disorders: reproductive life cycle factors". Advances in Nursing Science. 28 (4): 364–75. doi:10.1097/00012272-200510000-00008. PMID16292022. S2CID9172877.
^Lasiuk GC, Hegadoren KM (October 2007). "The effects of estradiol on central serotonergic systems and its relationship to mood in women". Biological Research for Nursing. 9 (2): 147–60. doi:10.1177/1099800407305600. PMID17909167. S2CID37965502.
^Sreeja S, Santhosh Kumar TR, Lakshmi BS, Sreeja S (July 2012). "Pomegranate extract demonstrate a selective estrogen receptor modulator profile in human tumor cell lines and in vivo models of estrogen deprivation". The Journal of Nutritional Biochemistry. 23 (7): 725–32. doi:10.1016/j.jnutbio.2011.03.015. PMID21839626.
^Thomas CG, Strom A, Lindberg K, Gustafsson JA (June 2011). "Estrogen receptor beta decreases survival of p53-defective cancer cells after DNA damage by impairing G₂/M checkpoint signaling". Breast Cancer Research and Treatment. 127 (2): 417–27. doi:10.1007/s10549-010-1011-z. PMID20623183. S2CID6752694.
^Collins P, Rosano GM, Sarrel PM, Ulrich L, Adamopoulos S, Beale CM, McNeill JG, Poole-Wilson PA (July 1995). "17 beta-Estradiol attenuates acetylcholine-induced coronary arterial constriction in women but not men with coronary heart disease". Circulation. 92 (1): 24–30. doi:10.1161/01.CIR.92.1.24. PMID7788912.
^Soltysik K, Czekaj P (April 2013). "Membrane estrogen receptors - is it an alternative way of estrogen action?". Journal of Physiology and Pharmacology. 64 (2): 129–42. PMID23756388.
^Boron WF, Boulpaep EL (2003). Medical Physiology: A Cellular And Molecular Approach. Elsevier/Saunders. p. 1300. ISBN978-1-4160-2328-9.
^Mutschler E, Schäfer-Korting M (2001). Arzneimittelwirkungen (in German) (8 ed.). Stuttgart: Wissenschaftliche Verlagsgesellschaft. pp. 434, 444. ISBN978-3-8047-1763-3.
^Wu CH, Motohashi T, Abdel-Rahman HA, Flickinger GL, Mikhail G (August 1976). "Free and protein-bound plasma estradiol-17 beta during the menstrual cycle". The Journal of Clinical Endocrinology and Metabolism. 43 (2): 436–45. doi:10.1210/jcem-43-2-436. PMID950372.
^Cheng ZN, Shu Y, Liu ZQ, Wang LS, Ou-Yang DS, Zhou HH (February 2001). "Role of cytochrome P450 in estradiol metabolism in vitro". Acta Pharmacologica Sinica. 22 (2): 148–54. PMID11741520.
^ abDorfman, Ralph I. (1961). "Steroid Hormone Metabolism". Radioactive Isotopes in Physiology Diagnostics and Therapy / Künstliche Radioaktive Isotope in Physiologie Diagnostik und Therapie. pp. 1223–1241. doi:10.1007/978-3-642-49761-2_39. ISBN978-3-642-49477-2.
^ ab"Estradiol"(PDF). ilexmedical.com. Archived(PDF) from the original on 4 February 2024. Retrieved 4 July 2024.
^ abcdeBecker JB, Berkley KJ, Geary N, Hampson E, Herman JP, Young E (4 December 2007). Sex Differences in the Brain: From Genes to Behavior. Oxford University Press. pp. 64–. ISBN978-0-19-804255-6. Estradiol levels are minimal during the earliest days of the follicular phase, but increasing concentrations are released into the general circulation as the follicle matures. The highest levels are reached about 24 to 48 hours before the LH peak. In fact, the pre-ovulatory peak in estradiol represents its highest concentration during the entire menstrual cycle. Serum concentrations at this time are typically about 130–200 pg/mL, but concentrations as high as 300–400 pg/mL can be achieved in some women. Following a transient fall in association with ovulation, estradiol secretion is restored by production from the corpus luteum during the luteal phase. Plateau levels of around 100–150 pg/mL (Abraham, 1978; Thorneycroft et al., 1971) are most often seen during the period from −10 to −5 days before the onset of menses. With the regression of the corpus luteum, estradiol levels fall, gradually in some women and precipitously in others, during the last few days of the luteal phase. This ushers in the onset of menses, the sloughing of the endometrium. Serum estradiol during menses is approximately 30–50 pg/mL. (Source.)
^ abcStrauss JR, Barbieri RL (2009). Yen and Jaffe's Reproductive Endocrinology: Physiology, Pathophysiology, and Clinical Management. Elsevier Health Sciences. pp. 807–. ISBN978-1-4160-4907-4. Archived from the original on 10 January 2023. Retrieved 21 December 2016. In most laboratories, serum estradiol levels range from 20 to 80 pg/mL during the early to midfollicular phase of the menstrual cycle and peak at 200 to 500 pg/mL during the preovulatory surge. During the midluteal phase, serum estradiol levels range from 60 to 200 pg/mL.
^ abcChristian C, von Schoultz B (15 March 1994). Hormone Replacement Therapy: Standardized or Individually Adapted Doses?. CRC Press. pp. 60–. ISBN978-1-85070-545-1. Plasma levels of estradiol range from 40 to 80 pg/mL during the 1st week of the ovarian cycle (early follicular phase) and from 80 to 300 pg/mL during the 2nd week (mid- and late follicular phase including periovulatory peak). Then during the 3rd and 4th weeks, estradiol fluctuates between 100 and 150 pg/mL (early and mid-luteal phase) to 40 pg/mL a few days before menstruation (late luteal phase). The mean integrated estradiol level during a full 28-day normal cycle is around 80 pg/mL.
^ abMüller EE, MacLeod RM (6 December 2012). Neuroendocrine Perspectives. Springer Science & Business Media. pp. 121–. ISBN978-1-4612-3554-5. Archived from the original on 10 January 2023. Retrieved 22 October 2016. [...] [premenopausal] mean [estradiol] concentration of 150 pg/mL [...]
^Powers MS, Schenkel L, Darley PE, Good WR, Balestra JC, Place VA (August 1985). "Pharmacokinetics and pharmacodynamics of transdermal dosage forms of 17 beta-estradiol: comparison with conventional oral estrogens used for hormone replacement". Am. J. Obstet. Gynecol. 152 (8): 1099–106. doi:10.1016/0002-9378(85)90569-1. PMID2992279.
^Kuhl H (2003). "Östrogene für den Mann?" [Estrogens for the man?]. Blickpunkt der Mann. 1 (3): 6–12. ISSN1727-0669. Archived from the original on 22 August 2023. Retrieved 22 August 2023.
^ abValues taken from day 1 after LH surge in: Stricker R, Eberhart R, Chevailler MC, Quinn FA, Bischof P, Stricker R (2006). "Establishment of detailed reference values for luteinizing hormone, follicle stimulating hormone, estradiol, and progesterone during different phases of the menstrual cycle on the Abbott ARCHITECT analyzer". Clinical Chemistry and Laboratory Medicine. 44 (7): 883–7. doi:10.1515/CCLM.2006.160. PMID16776638. S2CID524952. as PDF
^ abcdTotal amount multiplied by 0.022 according to 2.2% presented in: Wu CH, Motohashi T, Abdel-Rahman HA, Flickinger GL, Mikhail G (August 1976). "Free and protein-bound plasma estradiol-17 beta during the menstrual cycle". The Journal of Clinical Endocrinology and Metabolism. 43 (2): 436–45. doi:10.1210/jcem-43-2-436. PMID950372.[original research?]
^Coenjaerts M, Pape F, Santoso V, Grau F, Stoffel-Wagner B, Philipsen A, Schultz J, Hurlemann R, Scheele D (September 2021). "Sex differences in economic decision-making: Exogenous estradiol has opposing effects on fairness framing in women and men". Eur. Neuropsychopharmacol. 50 (2): 46–54. doi:10.1016/j.euroneuro.2021.04.006. hdl:20.500.11811/11066. ISSN0924-977X. PMID33957337. S2CID233982738.
^Newerla GJ (1944). "The History of the Discovery and Isolation of the Female Sex Hormones". New England Journal of Medicine. 230 (20): 595–604. doi:10.1056/NEJM194405182302001. ISSN0028-4793.
^Chemicals Identified in Human Biological Media: A Data Base. Design and Development Branch, Survey and Analysis Division, Office of Program Integration and Information, Office of Pesticides and Toxic Substances, Environmental Protection Agency. 1981. pp. 114–.